Pour les chiroptères, la forêt revêt une importance capitale, car c’est là qu’ils trouvent nourriture, habitats et protection. Afin d’en apprendre davantage sur le comportement nocturne des chiroptères dans la forêt, nous avons combiné de nouvelles méthodes de reconnaissance automatique des espèces avec des mesures détaillées de la structure de la forêt. Dans ce but, nous avons ajouté aux relevés d’habitat sur le terrain, des données de télédétection par laser (Light Detection and Ranging, LiDAR). LiDAR fournit de nouvelles informations sur le modèle d’activité des chiroptères auxquelles nous n’avons pas accès avec les relevés d’habitats ordinaires sur le terrain. Les données LiDAR sont prélevées sur une surface toujours plus étendue et constituen donc une source d’informations précieuse, qui jouera un rôle croissant pour la recherche sur la biodiversité en forêt.
Introduction
Presque un tiers de la surface de la Suisse est couverte de forêt. Cet écosystème est utilisé jour et nuit par un grand nombre d’organismes, parmi lesquels les chiroptères. À ce jour, trente espèces de chiroptères ont été identifiées en Suisse, ce qui représente un tiers de la diversité des espèces mammifères.
Les chiroptères sont uniques à maints égards: ce sont les seuls mammifères capables de voler; ils sont actifs la nuit; ils chassent et s’orientent dans l’obscurité par écholocation aux ultra-sons. Ils passent les jours froids et l’hiver en hibernation et peuvent vivre jusqu’à quarante ans. Toutes les espèces de chiroptères indigènes sont insectivores, et apportent ainsi une contribution écologique importante en réduisant le nombre de nuisibles, hétérocères et mouches nocturnes. Actifs la nuit, les chiroptères vont et viennent de leur quartier de jour à leur terrain de chasse. Chez presque toutes les espèces, l’un des deux se situe en forêt. Ainsi les murins de Daubenton (Myotis daubentonii) passent le plus clair de la journée dans les arbres creux de la forêt et, la nuit tombée, ils partent chasser les insectes sur les rives ou à la surface des plans d’eau en milieu ouvert. À l’inverse les Grands murins (Myotis myotis) élèvent leur progéniture en zone urbaine, gîtant dans les combles, mais préfèrent chasser en forêt, par exemple des carabes.
La forêt offre une grande diversité de structures qui sont diversement utilisées par les chauves-souris: arbres creux comme gîtes; forêt de feuillus pour se nourrir de carabes; structures de lisières intérieures et extérieures pour chasser les insectes croissant sur la végétation; petites branches dégagées pour la chasse à l’affût de papillons nocturnes de passage.

Ces diverses niches sont utilisées par différentes espèces qui se sont adaptées très différemment au cours de l’évolution en termes d’anatomie et de comportement. Ainsi la Noctule commune (Nyctalus noctula), avec ses ailes longues et étroites, chasse en vol rapide au-dessus des bois et repère ses proies sur de grandes distances avec des cris d’orientation de fréquence basse (20 kHz). Au contraire, le Petit rhinolophe (Rhinolophus hipposideros) émet des ultrasons à une fréquence très élevée (110 kHz), dont il capte la réflexion pour écholocaliser les battements d’ailes d’insectes nocturnes de passage afin de pouvoir les «cueillir» sur la végétation dans un vol très agile. Un grand groupe d’espèces, dont fait partie la chauve-souris la plus répandue en Suisse, la Pipistrelle commune (Pipistrellus pipistrellus), est moins spécialisé et parvient à se nourrir de façon opportuniste, notamment en se servant dans les «concentrations de nourriture» créées par l’activité humaine, par exemple autour des lampadaires.
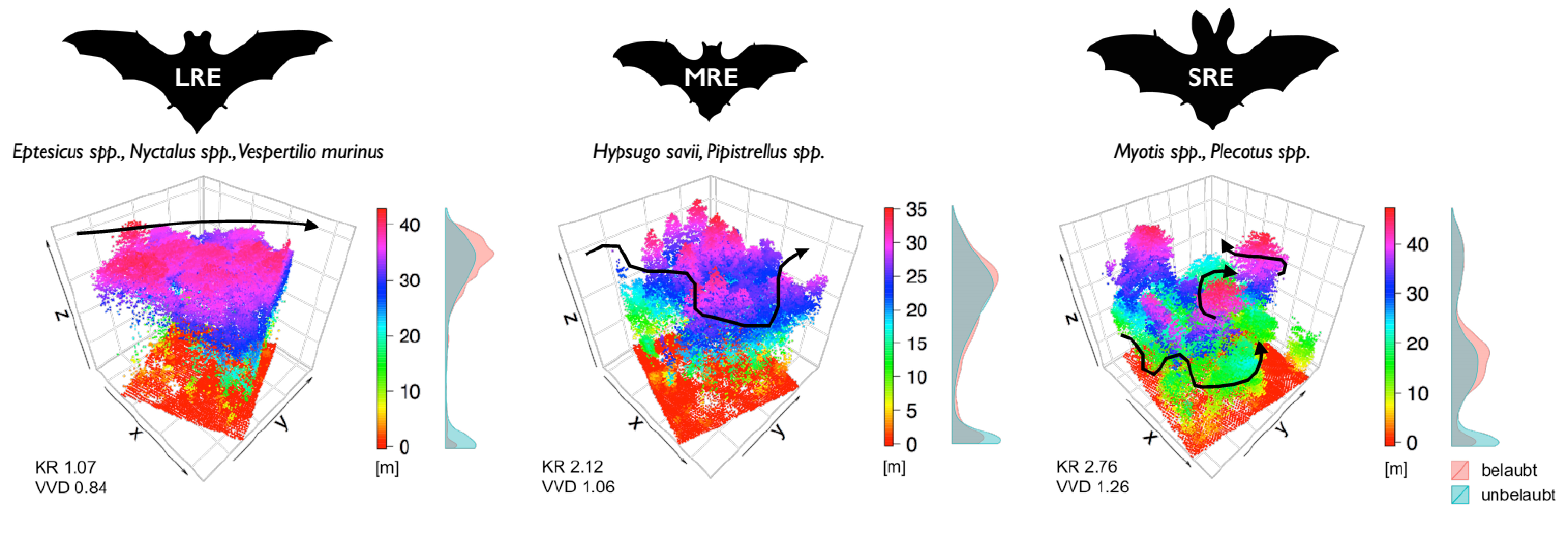
Symétriquement aux trois exemples cités, on peut répartir les espèces indigènes en trois guildes: chasseurs rapides en terrain ouvert, écholocalisant à longue distance (long range echolocators = LRE); chasseurs lents et agiles en vol, écholocalisant à courte distance (short range echolocators = SRE); enfin les chasseurs généralistes en structures de bordure, écholocalisant à distance moyenne (mid range echolocators = MRE). La technologie moderne, en l’occurrence l’écholocalisation des animaux, nous ouvre une fenêtre d’observation indirecte: des enregistreurs d’ultrasons (Batlogger) permettent l’enregistrement de leur activité acoustique et le logiciel développé par le WSL (BatScope), de déterminer l’espèce d’abord, puis son appartenance à l’une des guildes.
La question qui nous intéressait était de savoir si la structure forestière influence le comportement des chiroptères et quelles méthodes de mesure de cette structure sont les mieux indiquées pour décrire cette relation. Les méthodes ordinaires de relevé de terrain utilisées pour mesurer la structure forestière peuvent, certes, fournir de précieuses informations sur la forêt, mais elles sont chères et se limitent à de petites surfaces d’échantillonnage. De plus, la canopée reste largement inaccessible sur le terrain, alors qu’elle est une partie essentielle et importante au plan fonctionnel. Les nouvelles méthodes fondées sur la télédétection s’avèrent très utiles en l’espèce. Une méthode très prometteuse se base sur les informations que l’on peut recueillir par télédétection par laser (Light Detection and Ranging; LiDAR). Les données LiDAR fournissent des images fiables, en haute résolution, sur la structure tridimensionnelle de la forêt et sont prélevées sur des surfaces toujours plus étendues (Morsdorf 2011). Notre étude avait pour but de déterminer dans quelle mesure les données LiDAR et les relevés de terrain sont appropriés pour expliquer le modèle d’activité des chiroptères dans la forêt.
Étude de cas dans le canton d’Argovie
En été 2013 nous avons enregistré pendant plusieurs nuit les cris des chiroptères sur huit kilomètres carrés dans le canton d’Argovie, et ce sur quatre sites différents. Afin d’obtenir l’échantillonnage le plus complet de la structure verticale et horizontale de la forêt, nous avons placé plusieurs détecteurs à l’intérieur de la forêt (un détecteur au sol et un dans la canopée) et dans les clairières. Au moyen du logiciel BatScope, nous avons ensuite pu mesurer efficacement l’énorme quantité de cris des chauves-souris (données approximatives) et calculer le taux d’activité par guilde.
Parallèlement nous avons effectué sur chacun des sites des relevés de terrain complets relatifs à la structure forestière. L’objectif était de décrire de la manière la plus complète possible la structure de la forêt en termes de préférences des chiroptères. Nous avons ainsi calculé la densité de la végétation du sous-bois, car une forte densité entrave aussi bien l’écholocalisation des chiroptères, qu’elle limite leur capacité à manœuvrer en vol. De plus nous avons mesuré la densité verticale ou calculé l’indice de superficie foliaire au moyen de photos hémisphériques. Les relevés de terrain sont abondamment décrits dans Froidevaux et al. (2015, 2016).
Tout en poursuivant le même but que pour les relevés de terrain, nous avons en plus mesuré d’importantes structures forestières. Dans ce but nous disposions de deux jeux de données territoriales LiDAR que le canton d’Argovie avait acquis en 2014. Il s’agissait d’un relevé d’été, où les branches sont feuillues, et d’un relevé d’hiver, où les arbres sont dénudés (Fig. 2). Ces données nous ont permis de mesurer par exemple la rugosité de la canopée. Nous avons postulé que certaines espèces de chiroptères profitent d’une rugosité élevée, car ce type de structure offre des conditions microclimatiques favorables pour les insectes (nourriture) et une protection. De plus, les nuages de points LiDAR (Fig. 2) nous fournissent des données détaillées sur la répartition verticale de la végétation dans la forêt. Nous avons utilisé ces données pour calculer la densité et la régularité la végétation sur le profil vertical d’une forêt, car nous nous attendions à trouver ici aussi un rapport entre l’écholocation et la capacité de manœuvrer des chiroptères.
Les chiroptères se partagent la forêt
Nos analyses montrent qu’il existe un rapport étroit entre l’activité spécifique à une guilde de chiroptères et la structure forestière. Ainsi, l’activité de la guilde MRE croît en même temps que la rugosité de la canopée, c’est-à-dire lorsque la hauteur de la couronne des arbres est irrégulière. Ces espèces spécialisées dans les surfaces de lisières verticales et horizontales profitent d’une hétérogénéité élevée de la canopée, car elles y trouvent, d’une part, un abri contre les prédateurs et le mauvais temps et, d’autre part, une nourriture plus abondante car les insectes y bénéficient de conditions microclimatiques favorables. Par ailleurs, nos évaluations ont montré que l’activité de la guilde MRE baisse avec l’augmentation de la densité verticale de la végétation. Ce rapport est facile à comprendre si l’on songe que les chiroptères appartenant à la guilde MRE ne sont pas adaptés de façon optimale pour chasser à l’intérieur d’une forêt à la végétation dense. Quant aux chiroptères de la guilde SRE, avec leur grande agilité, ils seraient bien capables de se débrouiller à l’intérieur d’une forêt, pourtant leur activité baisse aussi légèrement avec l’augmentation de la densité verticale de la végétation. Il semble que les espèces de la guilde SRE utilisent l’intérieur de la forêt jusqu’à un certain degré de densité, mais qu’elles évitent les forêts au profil trop dense. Jusque-là nos résultats montrent clairement comment les espèces de chiroptères qui ont évolué différemment se répartissent les niches disponibles. Mais ils montrent aussi que les niches peuvent être inaccessibles du fait de structures trop denses.
LiDAR offre de nouvelles perspectives
La comparaison entre les données relevées sur le terrain et les données LiDAR relatives à la structure de la forêt montre que LiDAR est mieux indiqué pour expliquer le rapport entre structure de la forêt et activité des chiroptères. C’est avant tout la combinaison des données LiDAR relevées pendant les phases végétative et non végétative qui représente le mieux la répartition tridimensionnelle de la végétation dans la forêt. Ainsi, nous avons pu mesurer la structure de la canopée de manière détaillée, ce qui n’est pas possible avec les méthodes ordinaires sur le terrain. LiDAR offre donc de nouvelles perspectives pour l’étude de la relation entre la structure forestière et les habitats des chiroptères. Un grand avantage de la méthode LiDAR tient au fait que les données fournissent non seulement des mesures exactes de la structure forestières, mais que celles-ci s’étendent à tout un territoire. Ainsi, les futurs survols enregistrés par LiDAR nous permettront de quantifier efficacement toute modification de la structure de la forêt et des propriétés importantes des habitats, et ce sur toute l’étendue d’un vaste territoire. LiDAR représente aussi une source d’informations complète qui offre, surtout dans des écosystèmes tridimensionnels et complexes comme la forêt, de nombreuses possibilités d’application pour la recherche sur la protection de la nature. LiDAR ouvre également de nouvelles perspectives dans la compréhension du rapport entre structure forestière et biodiversité.
Remerciements
Nos remerciements vont à la Division Forêts du canton d’Argovie qui nous a permis de mener à bien la présente étude et qui nous a donné la possibilité d’utiliser les données cantonales LiDAR. Nous remercions également la Fondation pour la protection des chiroptères qui a mis à notre disposition les silhouettes de chauves-souris pour la figure 2. La thèse de Florian Zellweger a reçu le soutien du Fonds national suisse pour la recherche scientifique (numéro de projet 146786).
Bibliographie
- Froidevaux JSP, Zellweger F, Bollmann K, Obrist MK, 2015. Élaborer un plan d’échantillonnage acoustique fiable avec les logiciels «PRESENCE» et «GENPRES». Using “PRESENCE” and “GENPRES” software for designing a reliable acoustic sampling of bats. Le Vespère 5: 333-347.
- Froidevaux JSP, Zellweger F, Bollmann K, Jones G, Obrist MK, 2016. From field surveys to LiDAR: Shining a light on how bats respond to forest structure. Remote Sensing of Environment 175: 242-250.
- Morsdorf F, 2011. Erfassung struktureller Waldparameter mithilfe von flugzeuggetragenem Laserscanning. Journal forestier suisse 162: 164-170.
Interlocuteurs
Florian Zellweger1,2
Jérémy Froidevaux1,3
Kurt Bollmann1
Martin K. Obrist1
1 Institut fédéral de recherches WSL, 8903 Birmensdorf
2 Waldökologie, ETH Zürich, 8092 Zürich
3 University of Bristol, Bristol BS8 1TH, UK